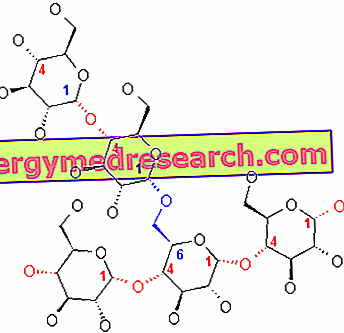
El glucógeno es una macromolécula (masa molecular de aproximadamente 400 millones de daltons) de α-glucosa en la que existen principalmente enlaces glicosídicos α-1, 4 y una ramificación en una proporción de 1:10, debido a los enlaces glicosídicos α-1, 6.
El glucógeno es un material de reserva y se degrada y reconstituye continuamente; En toda la masa celular del cuerpo, hay aproximadamente 100 g de glucógeno: la mayor parte se encuentra en el hígado donde es móvil y, por lo tanto, puede utilizarse como reserva para los otros órganos (el glucógeno en los músculos no es móvil).
Las enzimas que catalizan la degradación y la síntesis de glucógeno están todas en el citoplasma, por lo que necesitamos un sistema de ajuste que haga que una inactiva esté inactiva cuando la otra está activa: si hay disponibilidad de glucosa, esta última se convierte en glucógeno. (anabolismo), que es una reserva, y viceversa, si hay una demanda de glucosa, el glucógeno se degrada (catabolismo).
La enzima involucrada principalmente en la degradación del glucógeno es la glucógeno fosforilasa ; esta enzima es capaz de romper un enlace glicosídico α-1, 4 utilizando, como agente lítico, un ortofosfato inorgánico: la división tiene lugar por vía fosforolítica y se obtiene glucosa 1-fosfato.
A cinco o seis unidades de un punto de ramificación, la enzima glucógeno fosforilasa ya no puede actuar, por lo tanto, se separa del glucógeno y es reemplazada por una enzima desramificadora que es una transferasa : en el sitio catalítico de esta enzima hay una histidina que permite la transferencia de tres unidades de sacáridos a la cadena de glucósido más cercana (la histidina ataca al primer carbono de una molécula de glucosa). La enzima que acabamos de mencionar es la glicosiltransferasa ; Al final de la acción de esta enzima, solo queda una unidad de glucosa con el primer carbono unido al sexto carbono de una glucosa de la cadena principal. La última unidad de glucosa de la cadena lateral se libera por la acción de la enzima α-1, 6 glicosidasa (esta enzima constituye la segunda parte del desamificante enzimático); Dado que las ramificaciones en el glucógeno están en una proporción de 1:10, a partir de la degradación completa de la macromolécula obtenemos aproximadamente el 90% de glucosa 1-fosfato y aproximadamente el 10% de glucosa.
La acción de las enzimas mencionadas anteriormente permite eliminar una cadena lateral de la molécula de glucógeno; La actividad de estas enzimas se puede repetir hasta que se produzca una degradación completa de la cadena.
Considere un hepatocito; La glucosa (asimilada a través de la dieta), cuando ingresa a la célula se convierte en glucosa 6-fosfato y así se activa. La glucosa 6-fosfato, debido a la acción de la fosfoglucomutasa, se transforma en glucosa 1-fosfato: este último es un precursor no inmediato de la biosíntesis; En la biosíntesis se usa una forma activada de azúcares representada por el azúcar unido a un difosfato: generalmente uridildifosfato (UDP). La glucosa 1-fosfato se convierte luego en UDP-glucosa; este metabolito se somete a la acción de la glucógeno sintasa, que es capaz de unir UDP-glucosa a un extremo no reductor del glucógeno en crecimiento: se obtiene el glucógeno alargado de una unidad de glucósido y UDP. UDP es convertido por la enzima nucleosidedifosfochinasi en UTP que vuelve a la circulación.
La degradación del glucógeno se produce por la acción de la glucógeno fosforilasa que libera una molécula de glucosa y la transforma en glucosa 1-fosfato. Posteriormente, la fosfoglucomutasa convierte la glucosa 1-fosfato en glucosa 6-fosfato.
El glucógeno se sintetiza, sobre todo, en el hígado y en los músculos: en el organismo hay 1-1.2 hectolitros de glucógeno distribuidos por toda la masa muscular.
El glucógeno de un miocito representa una reserva de energía solo para esa célula, mientras que el glucógeno contenido en el hígado también es una reserva para otros tejidos, es decir, se puede enviar, como la glucosa, a otras células.
La glucosa 6-fosfato obtenida en los músculos a partir de la degradación del glucógeno se envía, en caso de requerimientos de energía, a la glucólisis; en el hígado, la glucosa 6-fosfato se convierte en glucosa por la acción de la glucosa 6-fosfato fosfatasa (enzima característica de los hepatocitos) y se canaliza hacia la corriente circulatoria.
La glucógeno sintasa y la glucógeno fosforilasa funcionan tanto en unidades de glucógeno no reductoras, por lo que debe haber una señal hormonal que ordene la activación de una ruta y la otra (o viceversa).
En el laboratorio fue posible alargar la cadena de glucógeno, explotando la glucógeno fosforilasa y utilizando glucosa 1-fosfato en una concentración muy alta.
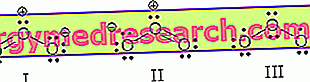
En las células, la glucógeno fosforilasa cataliza solo la reacción de degradación porque las concentraciones de los metabolitos son tales como para desplazar el balance de la siguiente reacción hacia la derecha (es decir, hacia la degradación del glucógeno):

Vemos el mecanismo de acción de la glucógeno fosforilasa: el oxígeno del acetal (que actúa como un puente entre las unidades de glucosa) se une al hidrógeno de fosforilo: se forma una reacción intermedia dada por un carbocatione (en la glucosa que es extremidad) a la que el fosforilo (Pi) se une muy rápidamente.
La glucógeno fosforilasa requiere un cofactor que es piridoxal fosfato (esta molécula también es un cofactor para las transaminasas): solo tiene protones parcialmente fosforilados (el piridoxal fosfato está rodeado por un ambiente hidrofóbico que justifica la presencia de protones unidos a él) . El fosforilo (Pi) es capaz de producir un protón para el glucógeno porque este fosforilo vuelve a adquirir el protón del fosforilo parcialmente protegido del fosfato de piridoxal. La probabilidad de que, a un pH fisiológico, el fosforilo pierda el protón y permanezca completamente desprotonada es muy baja.
Veamos ahora cómo funciona la fosfoglucomutasa. Esta enzima tiene un residuo de serina fosforilada en el sitio catalítico; la serina produce fosforilo en glucosa 1-fosfato (en la posición seis): durante un breve período de tiempo, la glucosa se forma en 1, 6-bisfosfato, luego la serina se refrigera al tomar el fosforilo en la posición uno. El phosphoglucus mutase puede funcionar en ambas direcciones, es decir, convertir la glucosa 1-fosfato en glucosa 6-fosfato o viceversa; Si se produce 6-fosfato de glucosa, puede enviarse directamente a la glucólisis, a los músculos o transformarse en glucosa en el hígado.
La enzima uridil fosfoglucus transferasa (o UDP glucosa pirofosforilasa) cataliza la reacción de transferencia de glucosa-1-fosfato a la UTP al atacar la fosforila a.
La enzima descrita anteriormente es una pirofosforilasa: este nombre se debe al hecho de que la reacción opuesta a la que acabamos de describir es la pirofosforilación.
La glucosa UDP, obtenida como se describe, puede alargar la cadena de glucógeno de una unidad de monosacárido.
Es posible evolucionar la reacción hacia la formación de glucosa UDP mediante la eliminación de un producto que es el pirofosfato; La enzima pirofosfatasa convierte el pirofosfato en dos moléculas de ortofosfato (hidrólisis de un anhídrido) y al hacerlo, mantiene la concentración de pirofosfato tan baja que el proceso de formación de glucosa UDP se promueve termodinámicamente.
Como se mencionó, la glucosa UDP, gracias a la acción de la glucógeno sintasa, es capaz de alargar la cadena de glucógeno.
Las ramificaciones (en una proporción de 1:10) se deben al hecho de que, cuando una cadena de glucógeno está formada por 20-25 unidades, interviene una enzima ramificada (que tiene una istidina en su sitio catalítico), capaz de transferir una serie de 7 -8 unidades glicosídicas aguas abajo de 5-6 unidades: así se genera una nueva rama.
Para cuestiones de origen nervioso o si se requiere energía debido al esfuerzo físico, la adrenalina se secreta de las glándulas suprarrenales.
Las células diana de la adrenalina (y la norepinefrina) son las del hígado, los músculos y el tejido adiposo (en este último existe la degradación de los triglicéridos y la circulación de los ácidos grasos: en la mitocondria se produce, por consiguiente, glucosa 6 -fosfato, que se envía a la glucólisis, mientras que en los adipocitos, la glucosa 6-fosfato se transforma en glucosa por la enzima glucosa 6-fosfato fosfatasa y se exporta a los tejidos).
A ver, ahora los modos de acción de la adrenalina. La adrenalina se une a un receptor ubicado en la membrana celular (de los miocitos y los hepatocitos) y esto determina la traducción de la señal desde el exterior hacia el interior de la célula. La proteína quinasa se activa y actúa simultáneamente en los sistemas que regulan la síntesis y degradación del glucógeno:
La glucógeno sintasa existe en dos formas: una forma desfosforilada (activa) y una forma fosforilada (inactiva); La proteína cinasa fosforila la glucógeno sintasa y bloquea su acción.
La glucógeno fosforilasa puede existir en dos formas: una forma activa en la que está presente una serina fosforilada y una forma inactiva en la que la serina está desfosforilada. La glucógeno fosforilasa puede ser activada por la enzima glucógeno fosforilasa quinasa . La glucógeno fosforilasa quinasa es activa si está fosforilada e inactiva si está desfosforilada; la proteína quinasa tiene como sustrato la glucógeno fosforilasa quinasa, es decir, es capaz de fosforilar (y, por lo tanto, activar) esta última que, a su vez, activa la glucógeno fosforilasa.
Una vez que la señal de adrenalina ha terminado, también debe terminar su efecto en la célula: las enzimas fosfatasa intervienen, entonces, sobre la especie de proteína.